Ahhoz, hogy a fehérje szerkezetét alkotó húsz aminosavról és a módosítottról beszéljünk, legalább tizenkét speciális metabolikus útvonalat kell leírni.
De miért használnak a sejtek olyan sok anyagcsere utat, amelyek energiát igényelnek (például az enzimek katalitikus helyeinek regenerálásához), mindegyik enzimatikus örökséggel, az aminosavak katabolizálásához? Szinte minden aminosavból speciális utakon keresztül olyan metabolitok nyerhetők, amelyek kis részben energiatermelésre szolgálnak (például glükoneogenezis és ketontestek útján), de mindenekelőtt komplexek kialakulásához vezetnek nagyszámú szénatomot tartalmazó molekulák (például fenilalaninból és tirozinból, hormonok termelődnek az erre a célra szánt mellékvesékben); ha egyrészt egyszerű lenne energiát előállítani aminosavakból, másrészt bonyolult lenne kis molekulákból kiindulva összetett molekulákat építeni: az aminosavak katabolizmusa lehetővé teszi számukra, hogy kihasználják csontvázukat, hogy nagyobb fajokat kapjanak.
Egy egészséges egyén naponta két vagy három hektogramm aminosavat bont le: 60-100 g abból az étrendben bevezetett fehérjékből származik, de több mint 2 hektogramot a szervezet szerves részét képező fehérjék normális forgalmából nyernek ( e fehérjék aminosavai, amelyeket a redox folyamatok károsítanak, másokkal helyettesítik és katabolizálják).
Az aminosavak energiával járulnak hozzá az ATP-hez: az α-aminocsoport eltávolítása után az aminosavak maradék széntartalmú váza megfelelő átalakításokat követően beléphet a krebs-ciklusba. Továbbá, ha a tápanyag -ellátottság hiányzik, és a glükóz mennyisége csökken, a glükoneogenezis aktiválódik: a glükoneogenetikus aminosavakról azt mondják, hogy megfelelő módosítások után bevihetők a glükoneogenezisbe; a glükoneogenetikus aminosavak azok, amelyek átalakíthatók piruvátban vagy fumarátban (a fumarát a mitokondriumot elhagyó maláttá alakítható, és a citoplazmában oxaloacetáttá alakul, amelyből foszfoenol-piruvát nyerhető). ecet-acetát.
Az imént leírt nagyon fontos szempont, mert az aminosavak orvosolhatják a cukorhiányt azonnali böjt esetén; ha az éhezés továbbra is fennáll, két nap elteltével a lipid anyagcsere beavatkozik (mivel a fehérjék szerkezetét nem lehet túlságosan megtámadni), ebben a fázisban, mivel a glükoneogenezis nagyon korlátozott, a zsírsavak acetil -koenzim A és ketontestekké alakulnak át. A további koplalástól kezdve az agy is alkalmazkodik a ketontestek használatához.
Az α-aminocsoport átvitele az aminosavakból transzaminációs reakción keresztül történik; az enzimek, amelyek katalizálják ezt a reakciót, azt mondják, valójában transzaminázok (vagy aminotranszferáz). Ezek az enzimek egy piridoxál -foszfát nevű enzimatikus kofaktort használnak, amely közbeavatkozik az aldehidcsoportjával. A piridoxál -foszfát a piridoxin foszforilációjának terméke, amely főleg zöldségekben található vitamin (B6).
A transzaminázok a következő tulajdonságokkal rendelkeznek:
Nagy specifitás a ketoglutarát-glutamát α-párra;
A második párról nevezték el őket.
A transzamináz enzimek mindig magukban foglalják az α ketoglutarát-glutamát párt, és a második érintett pár szerint különböznek egymástól.
Példák:
L "aszpartát transzamináz azaz GOT (Glutamate-Ossal acetate Transaminase): az enzim az α-amino-csoportot az aszpartátból az α-ketoglutarátba transzferálja, oxaloacetátot és glutamátot kapva.
L "alanin transzamináz azaz GTP (glutamát-piruvátát-transzamináz): az enzim átviszi az α-amino-csoportot az "alaninból" az "a-ketoglutarátba", így piruvátot és glutamátot kap.
A különböző transzaminázok az α-ketoglurátot használják az aminosavak aminocsoportjának elfogadójaként, és glutamáttá alakítják; míg a képződött aminosavakat a ketontestek útvonalában használják fel.
Ez a fajta reakció mindkét irányban megtörténhet, mivel megszakadnak és azonos energiatartalmú kötéseket hoznak létre.
A transzaminázok mind a citoplazmában, mind a mitokondriumban találhatók (többnyire a citoplazmában aktívak), és izoelektromos pontjukban különböznek egymástól.
A transzaminázok képesek dekarboxilezni az aminosavakat is.
Létre kell hozni a glutamát α-ketoglutaráttá való átalakításának módját: ezt dezaminálással végzik.
Ott glutamát -dehidrogenáz ez egy olyan enzim, amely képes a glutamátot α-ketoglutaráttá alakítani, és ezért a glutamát formájában található aminosavak aminocsoportjait ammóniává alakítani. Ez egy redox folyamat, amely áthalad az α-amino-glutarát köztes termékén: ammónia és α-ketoglutarát szabadul fel, és visszatér a keringésbe.
Ezután az aminosavak aminocsoportjainak ártalmatlanítása áthalad a transzaminázokon (amelyek szubsztráttól függően különböznek) és a glutamát -dehidrogenázon, amely meghatározza az ammónia képződését.
A glutamát -dehidrogenáznak két típusa létezik: citoplazmatikus és mitokondriális; a kofaktor, amely ezen enzim kozubsztrátja is, a NAD (P) +: a glutamát -dehidrogenáz vagy a NAD + -t, vagy a NADP + -ot használja a redukáló erő elfogadójaként. A citoplazmatikus forma előnyben részesíti - bár nem kizárólagosan - a NADP + -ot, míg a mitokondriális forma a NAD + -ot. A mitokondriális forma célja az aminocsoportok ártalmatlanítása: ammónia (amely a mitokondriumban található speciális enzim szubsztrátja) és NADH (amely a légzőláncba kerül) képződéséhez vezet. A citoplazmatikus forma az ellenkező irányban működik, azaz ammóniát és α-ketoglutarátot használ a glutamát előállítására (amelynek bioszintetikus célja van): ez a reakció egy reduktív bioszintézis, és az alkalmazott kofaktor a NADPH.
A glutamát -dehidrogenáz akkor működik, ha szükség van az aminosavak aminocsoportjainak (pl. Ammónia) ártalmatlanítására (vizelet útján), vagy ha az aminosavak csontvázai szükségesek az energia előállításához: ennek az enzimnek tehát negatív modulátorai lesznek a jelzőrendszerek jó energiaellátottságúak (ATP, GTP és NAD (P) H), és pozitív modulátorként az energiaigényt jelző rendszerek (AMP, ADP, GDP, NAD (P) +, aminosavak és pajzsmirigyhormonok).
Az aminosavak (főleg a leucin) a glutamát -dehidrogenáz pozitív modulátorai: ha aminosavak vannak a citoplazmában, felhasználhatók fehérjeszintézishez, vagy el kell dobni, mert nem halmozódhatnak fel (ez magyarázza, hogy az aminosavak miért pozitív modulátorok) ).
Az ammónia ártalmatlanítása: karbamid ciklus
A halak az ammóniát a kopoltyúkon keresztül vízbe juttatva, a madarak húgysavvá (kondenzációs termék) alakítják át, és ürülékkel távolítják el. Lássuk, mi történik az emberekben: azt mondtuk, hogy a glutamát-dehidrogenáz átalakítja a glutamátot α- ketoglutarátot és ammóniát, de nem mondtuk, hogy ez csak a máj mitokondriumában fordul elő.
Az ammónia elvezetésében alapvető szerepe van a karbamid cikluson keresztül a mitokondriális transzaminázoknak.

A szén-dioxidot hidrogén-karbonát ion (HCO3-) formájában aktiválja a biotin kofaktor, amely karboxi-biotint képez, amely ammóniával reagálva karbaminsavat képez; a következő reakcióban ATP-t használva foszfátot visz át a karbaminsavra, karbamil-foszfátot és ADP-t képezve (az ATP átalakítása ADP -vé a hajtóerő a karboxibiotin megszerzéséhez.) Ezt a fázist katalizálja karbamil -foszfát -szintetáz és a mitokondriumban fordul elő. A karbamil -foszfát és az ornitin az enzim szubsztrátjai ornitin transz -karbamiláz amely citrullinná alakítja őket; ez a reakció a mitokondriumokban (hepatociták) fordul elő. A keletkező citrullin elhagyja a mitokondriumot, és a citoplazmában a "hatás" alá kerülarginin -szukcinát szintetáz: van egy fúzió a citrullin és egy aszpartát széntartalmú váza között nukleofil támadással, majd a víz eltávolításával. Az arginin -szukcinát szintetáz enzimhez ATP -molekula szükséges, így energetikai kapcsolódás következik be: az ATP hidrolízise AMP -vé és pirofoszfáttá (ez utóbbit két ortofoszfát -molekulává alakítják át) egy molekula d "víz" kiürítésével történik. és nem a közeg vizének hatására.
A következő enzim aarginin -szukcináz: ez az enzim képes arginin -szukcinátot argininre és fumarátra bontani a citoplazmán belül.
A karbamid ciklust az enzim fejezi be argináz: karbamidot és ornitint kapunk; a karbamidot a vesék (vizelet) ártalmatlanítják, míg az ornitin visszatér a mitokondriumba és folytatja a ciklust.
A karbamid ciklust arginin közvetett modulációnak teszi ki: az arginin felhalmozódása azt jelzi, hogy a karbamid ciklust fel kell gyorsítani; az arginin modulációja közvetett, mivel az arginin pozitívan modulálja az acetil -glutamát szintetáz enzimet. Ez utóbbi acetilcsoportot képes átvinni a glutamát nitrogénjére: N-acetil-glutamát keletkezik, amely a karbamil-foszfo-szintetáz enzim közvetlen modulátora.
Az arginin a karbamidciklus metabolitjaként halmozódik fel, ha a karbamil -foszfát termelése nem elegendő az ornitin eltávolításához.
A karbamid csak a májban termelődik, de vannak olyan helyek, ahol a kezdeti reakciók lejátszódnak.
Az agy és az izmok speciális stratégiákat alkalmaznak az aminocsoportok kiküszöbölésére. Az agy nagyon hatékony módszert alkalmaz, amelyben enzimet használnak glutamin szintetáz és egy enzim glutamáz: az első a neuronokban van jelen, míg a második a májban. Ez a mechanizmus nagyon hatékony két okból:
Két aminocsoportot szállítanak az agyból a májba egyetlen hordozóval;
A glutamin sokkal kevésbé mérgező, mint a glutamát (a glutamát neuronátvitelt is végez, és nem lépheti túl a fiziológiai koncentrációt).
A halakban hasonló mechanizmus hozza az aminosavak aminocsoportját a kopoltyúkhoz.
Az izomzatból (csontváz és szív) az aminocsoportok a glükóz-alanin cikluson keresztül jutnak el a májba; az érintett enzim a glutamin-piruvát transzamináz: lehetővé teszi az aminocsoportok (amelyek glutamát formájában vannak) transzponálását, a piruvát alaninná történő átalakítását, és ezzel egyidejűleg a glutamát α-ketoglutaráttá alakítását az izomzatban, és katalizálja a fordított folyamat a májban.
A különböző feladatokkal vagy pozíciókkal rendelkező transzaminázok szerkezeti különbségekkel is rendelkeznek, és elektroforézissel meghatározhatók (különböző izoelektromos pontjaik vannak).
A transzaminázok vérben való jelenléte a máj- vagy szívkárosodás tünete lehet (azaz a máj vagy a szívsejtek szöveti károsodása); a transzaminázok nagyon magas koncentrációban vannak mind a májban, mind a szívben: elektroforézissel megállapítható, hogy a károsodás a májban vagy a szívsejtekben történt -e.
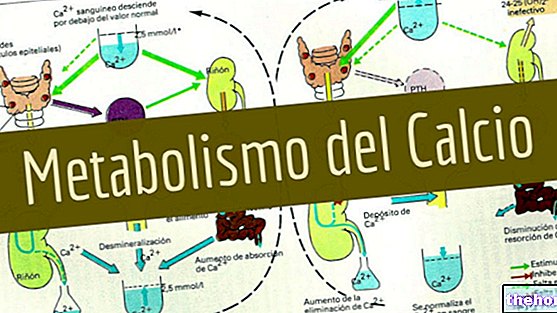
.jpg)


















---ena-screening-o-pannello-ena.jpg)
