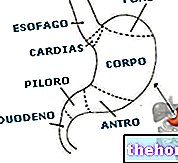
A fehérjék elemzése a következő átlagértékeket adja: 55% szén, 7% hidrogén és 16% nitrogén; egyértelmű, hogy a fehérjék különböznek egymástól, de átlagos elemi összetételük alig különbözik a fent megadott értékektől .
Alkotmányosan a fehérjék természetes α-aminosavakból képződő makromolekulák; az aminosavak az amidkötésen keresztül kapcsolódnak össze, amelyet egy a-aminosav aminocsoportja és egy másik a-aminosav karboxilja közötti reakció hoz létre.
Ezt a kötést (-CO-NH-) peptidkötésnek is nevezik, mivel peptideket (aminosavakat kombinálva) köt:
a kapott dipeptid, mivel két aminosavból áll. Mivel a dipeptid egyik végén szabad aminocsoportot (NH2), a másikban karboxilcsoportot (COOH) tartalmaz, így egy vagy több aminosavval reagálhat, és meghosszabbíthatja a láncot jobbról és balról is, ugyanazzal a reakcióval fentebb látva.
A reakciósorozat (ami egyébként nem is olyan egyszerű) a végtelenségig folytatódhat: amíg nem lesz polimer ún. polipeptid vagy fehérje. A peptidek és fehérjék megkülönböztetése a molekulatömeghez kapcsolódik: általában 10 000 -nél nagyobb molekulatömeg esetén fehérjékről beszélünk.
Az aminosavak összekötése még kis fehérjék előállításához is nehéz feladat, bár a közelmúltban kifejlesztettek egy automatikus módszert aminosavakból történő fehérjék előállítására, amely kiváló eredményeket ad.
A legegyszerűbb fehérje tehát 2 aminosavból áll: nemzetközi egyezmény szerint a fehérjeszerkezetben az aminosavak rendezett számozása a szabad a-aminocsoporttal rendelkező aminosavból indul ki.
kódolása), amely nem elhanyagolható kémiai nehézségekkel szembesül.
Az aminosavak rendezett szekvenciáját Edman-bomlás útján lehetett meghatározni: a fehérjét fenil-izotiocianáttal (FITC) reagáltatják, kezdetben az α-amino-nitrogén dublett megtámadja a fenil-izotiocianátot, amely a tiokarbamil-származékot képezi; ezt követően a kapott termék ciklizál, és fluoreszkáló fenil -tiohidantoin -származékot kap.
Edman kifejlesztett egy szekvenáló nevű gépet, amely automatikusan beállítja a paramétereket (idő, reagensek, pH stb.) A lebomlás érdekében, és biztosítja a fehérjék elsődleges szerkezetét (ezért Nobel -díjat kapott).
Az elsődleges szerkezet nem elegendő a fehérjemolekulák tulajdonságainak teljes értelmezéséhez; úgy vélik, hogy ezek a tulajdonságok lényegesen függenek attól a térbeli konfigurációtól, amelyet a fehérjemolekulák hajlamosak feltételezni, különböző módon hajtogatva: vagyis feltételezve azt, amit a fehérjék másodlagos szerkezetének definiáltak.
A fehérjék másodlagos szerkezete villódzik, vagyis hajlamos a felmelegedésre szétesni; akkor a fehérjék denaturálják magukat, sok jellemző tulajdonságukat elveszítve. A 70 ° C feletti hevítés mellett a denaturációt a besugárzás vagy a reagensek hatása is okozhatja (például erős savakból).
A fehérjék denaturálódása a termikus hatás miatt megfigyelhető például a tojásfehérje hevítésével: látható, hogy elveszíti kocsonyás megjelenését, és oldhatatlan fehér anyaggá alakul. A fehérjék denaturálása azonban másodlagos szerkezetük megsemmisüléséhez vezet, de változatlanul hagyja elsődleges szerkezetét (a különböző aminosavak összefűzését).
A fehérjék akkor veszik fel a harmadlagos szerkezetet, amikor a láncuk, bár a másodlagos szerkezet hajlítása ellenére még mindig rugalmas, de összehajlik, és így egy összehúzott, háromdimenziós elrendezést hoz létre szilárd test alakjában. A harmadlagos szerkezetért mindenekelőtt a diszulfidkötések felelősek, amelyek a molekula mentén szétszórt cisztein -SH között létesíthetők.
A negyedik szerkezet viszont csak a két vagy több alegység által alkotott fehérjékhez tartozik. A hemoglobin például két fehérjepárból (azaz mind a négy fehérje láncból) áll, amelyek egy tetraéder csúcsaiban helyezkednek el, hogy gömbszerű szerkezetet hozzanak létre; a négy fehérje láncot ionos és nem kovalens kötések.
Egy másik példa a negyedéves szerkezetre az inzulin, amely úgy tűnik, hogy akár hat fehérje alegységből áll, amelyek párban vannak elhelyezve egy háromszög csúcsán, amelynek közepén két cink atom található.
Szálas fehérjék
Ezek bizonyos merevségű és sokkal hosszabb tengelyű fehérjék, mint a másik; a természetben nagyobb mennyiségben jelen lévő rostos fehérje kollagén (vagy kollagén).
Egy rostos fehérje különböző másodlagos szerkezeteket feltételezhet: α-hélix, β-lemez és kollagén esetén hármas hélix; Az α-hélix a legstabilabb szerkezet, amelyet a β-lap követ, míg a három közül a legkevésbé stabil a hármas hélix.
jobb ha a fő csontvázat követve (alulról felfelé irányítva) a jobb oldali csavar csavarásához hasonló mozgást hajtanak végre; míg a hélix a bal kéz ha a mozgás analóg a bal oldali csavar csavarozásával. a -helikális kézzel az -R szubsztituensek befelé néznek. A jobbkezes a-hélixek stabilabbak, mint a balkezesek, mert a vati -R c "között kevesebb kölcsönhatás és kevesebb sztérikus akadály van. A fehérjékben található összes a-hélix dextrorotális.
Az α-hélix szerkezetét stabilizálják azok a hidrogénkötések (hidrogénhidak), amelyek az egyes aminosavak karboxilcsoportja (-C = O) és az aminosav (-NH) között keletkeznek lineáris sorrend.
Az α-hélix szerkezetű fehérje például a haj keratin.
Az α-hélix szerkezetének meghosszabbításával az α-hélixről a β-lemezre való átmenetet hajtjuk végre; a hő vagy a mechanikai igénybevétel is lehetővé teszi az α-hélixből a β-lemez szerkezetbe való átjutást.
Általában egy fehérjében a β-lapos szerkezetek közel vannak egymáshoz, mert láncközi hidrogénkötések hozhatók létre a fehérje részei között.
A rostos fehérjékben a fehérjeszerkezet nagy része α-hélixben vagy β-lapban szerveződik.
Globuláris fehérjék
Szinte gömb alakú térszerkezettel rendelkeznek (a polipeptidlánc irányának számos változása miatt); a lét egyes részei egy α-hélixre vagy β-lapos szerkezetre vezethetők vissza, és más részek nem tulajdoníthatók ezeknek a formáknak: az elrendezés nem véletlen, hanem rendszerezett és ismétlődő.
Az eddig említett fehérjék teljesen homogén szerkezetű anyagok: azaz kombinált aminosavak tiszta szekvenciái; ilyen fehérjéket mondanak egyszerű; vannak fehérjék, amelyek egy fehérje részből és egy nem fehérje részből (prosztata csoport) állnak, amelyeket fehérjéknek neveznek konjugált.
, a körmökben, a szaruhártyában és a szemlencsében, egyes szervek (pl. máj) intersticiális terei között stb.
Szerkezete különleges mechanikai képességeket biztosít; nagy mechanikai szilárdsággal jár, amely nagy rugalmassággal (pl. inakban) vagy nagy merevséggel (pl. csontokban) függ az elvégzendő funkciótól függően.
A kollagén egyik legérdekesebb tulajdonsága az alkotó egyszerűség: körülbelül 30% -ban prolinból és körülbelül 30% glicinből áll; a többi 18 aminosavnak csak a fehérje szerkezetének fennmaradó 40% -át kell megosztania. A kollagén aminosavszekvenciája feltűnően szabályos: minden három maradék esetében a harmadik a glicin.
A prolin egy ciklikus aminosav, amelyben az R csoport az α-amino-nitrogénhez kötődik, és ez bizonyos merevséget biztosít.
A végső szerkezet egy ismétlődő lánc, amelynek "hélix" alakja van; a kollagénláncban nincsenek hidrogénkötések. A kollagén egy "bal oldali hélix, amelynek dőlésszöge (hossza a spirál egy fordulatának felel meg) nagyobb, mint az" α-hélix; a kollagén-hélix annyira laza, hogy három fehérje lánc képes körbefogni egymást, és " egyetlen kötél: hármas spirál szerkezet.
A kollagén hármas hélix azonban kevésbé stabil, mint az α-hélix és a β-lapos szerkezet.
Nézzük most a kollagén előállításának mechanizmusát; Vegyük például egy véredény szakadását: ezt a szakadást számtalan jel kíséri azzal a céllal, hogy lezárja az edényt, és így létrejöjjön az alvadék.
Az alvadáshoz legalább harminc speciális enzim szükséges. Az alvadás után folytatni kell a szövet javítását; a sebhez közeli sejtek kollagént is termelnek. Ehhez először egy gén expresszióját indukáljuk, vagyis azokat a szervezeteket, amelyek egy gén információiból kiindulva képesek előállítani a fehérjét (a genetikai információ átíródik a mRNS -re, amely elhagyja a magot és eléri a riboszómákat. citoplazmában, ahol a genetikai információ fehérjévé alakul.) Ezután a kollagén szintetizálódik a riboszómákban (úgy néz ki, mint egy bal oldali hélix, amely körülbelül 1200 aminosavból áll, és molekulatömege körülbelül 150 000 d), majd felhalmozódik a lumenekben. szubsztrátja lesz azoknak az enzimeknek, amelyek képesek a poszt-transzlációs módosítások végrehajtására (a nyelv módosítása "mRNS-sel"); a kollagénben ezek a módosítások egyes oldalláncok, különösen a prolin és a lizin hidroxilezéséből állnak.
Az ezekhez a változásokhoz vezető enzimek kudarca skorbutot okoz: ez egy olyan betegség, amely kezdetben az erek törését, a fogak törését okozza, amelyet bélrendszeri vérzések és halál követhet; a hosszú élettartamú élelmiszerek folyamatos használata okozhatja.
Ezt követően más enzimek hatására más módosítások is előfordulnak, amelyek a prolin és a lizin hidroxilcsoportjainak glikozidálását tartalmazzák (egy cukor kötődik az OH oxigénjéhez); ezek az enzimek a lumenen kívül más területeken is megtalálhatók, ezért a fehérje, miközben módosul, az endoplazmatikus retikulumon belül vándorol, hogy zsákokba (vezikulákba) kerüljön, amelyek magukra záródnak és leválnak a retikulumról: bennük a glikozidált pro -kollagén monomer; az utóbbi eléri a Golgi készüléket, ahol bizonyos enzimek felismerik a glikozidált pro-kollagén karboxiterminális részében lévő ciszteint, és a különböző láncokat egymáshoz közelítik, és diszulfidhidakat képeznek: ily módon három lánc a glikozidált pro-kollagént összekapcsolva nyerik, és ez a kiindulópont, amelyből a három lánc egymásba hatolva spontán hármas hélixet hoz létre. elfojtja magát, leválasztja magát a Golgi -készülékről, és a három láncot a sejt perifériája felé szállítja, ahol a fúzión keresztül a plazmamembránnal, a trimeter kilökődik a sejtből.
Az extra sejtes térben vannak bizonyos enzimek, a pro-kollagén peptidázok, amelyek eltávolítják a sejtből kiűzött fajokból, három darab (mindegyik hélixből egy) 300 aminosav l-egy, a terminális karboxi-részből és három töredékek (egy -egy hélixhez), egyenként körülbelül 100 aminosavból, az aminoterminális részből: marad egy hármas hélix, amely hélixenként körülbelül 800 aminosavból áll. tropokollagén.
A tropokollagén meglehetősen merev rúdnak tűnik; a különböző trimmerek kovalens kötésekhez kapcsolódva nagyobb szerkezeteket eredményeznek: mikrofibrillák. A mikroszálakban a különböző trimerek lépcsőzetesen vannak elrendezve; sok mikrofibrillum tropokollagén köteget képez.
A csontokban, a kollagénrostok között intersticiális terek találhatók, amelyekben kalcium és magnézium szulfátjai és foszfátjai rakódnak le: ezek a sók az összes rostot is lefedik; ez merevvé teszi a csontokat.
Az inakban az intersticiális terek kevésbé gazdagok kristályokban, mint a csontok, míg kisebb fehérjék vannak jelen, mint a tropokollagénben: ez rugalmasságot kölcsönöz az inaknak.
A csontritkulás olyan betegség, amelyet a kalcium és a magnézium hiánya okoz, ami lehetetlenné teszi a sók rögzítését a tropokollagén rostok intersticiális területein.