EGYESÜLET (VAGY "CONCATENATION" VAGY "LINKAGE")
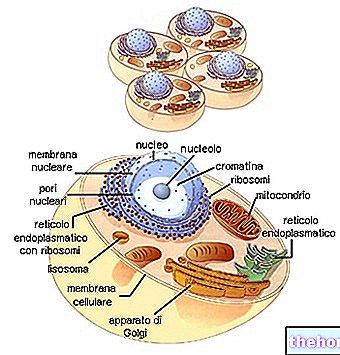
Eddig a mendeli dihibrid (vagy polihibrid) keresztezést úgy kezeltük, hogy feltételezzük, hogy a különböző allélpárok valójában különböző homológ kromoszómapárokon találhatók meg. elérheti a száz kromoszómát), míg a gének száma több tízezerre tehető.
Szerencsés esély volt, hogy a Mendel által kísérleteihez kiválasztott karakterek egymástól függetlenül elkülönültek (anélkül, hogy összezavarnák a számításokat a fenotípusok eloszlásában a polihibrid keresztezés F2 -ben). Szerencsés esély volt. követné, ezt társulási törvénynek hívnák.
Tudva, hogy nagyon sok karakternek egyetlen kromoszómán van a lókusza, és hogy azok a kromoszómapárok, amelyek egymástól függetlenül elkülönülnek a meiózisban, jól megérthető, hogy milyen gyakran fordul elő, hogy két pár karakter, ha a szülői kromoszómához kapcsolódnak egyformán az ivarsejtben, és ezért abban a szervezetben is, amelyhez saját genetikai anyagát hozza.
Így azt látjuk, hogy a "társulás" kivételt jelent, messze nem ritkán, a Mendel harmadik törvényében megfogalmazott függetlenség alól.
CSERÉLÉS VAGY "ÁTLÉPÉS" ÉS REKOMBINÁCIÓ
A meiózisról szólva jeleztük, hogy a genetikai anyag keveredésének két különböző pillanata van: az egyik az ivarsejtekben lévő kromoszómák szegregációja, és ezt Mendel is megfigyelte.
A másik pillanat, amely valójában megelőzi, az a pillanat, amelyben a homológ kromoszómapárok minden párjának négy kromatidája kölcsönösen azonos tulajdonságokat cserél. Ezt a cserét követően két, ugyanazon kromoszómához társított tényező független lesz az ivarsejtekben. hogy a csere zajlik, az első közelítéssel arányos a kromoszóma hosszával, és a hosszabb kromoszómákban akár több is lehet, mint a csere.
A jelenség citológiailag kimutatható, ha elegendő számú meiózist figyelünk meg mikroszkóp alatt.
Az újratelepítési arány az az arány, amellyel a szülői generációban társított két karakter eltérő módon rekombinálódik az F2-ben.
Ha a két lókusz teljesen szomszédos, akkor annak valószínűsége, hogy egy chiasma elválasztja őket, gyakorlatilag nulla. A rekombinációs ráta a következő lesz: n ° rekombináns. Ha két lókusz két különböző kromoszómán van, a rekombinációs ráta 0,5 lesz (egyenlő valószínűséggel, ha a P generációban összekapcsolt két karakter véletlenszerűen találja magát az F2 -ben). A rekombinációs ráta ezért 0,0 és 0,5 között változhat. A kromoszómán lévő kis távolságok esetén a távolság és a rekombinációs arány egyenesen arányos. Hosszabb távolságok esetén fennáll annak a lehetősége, hogy két csere történik két lókusz között. Most nyilvánvalóvá válik, hogy két, két cserével elválasztott tényező ismét összefügg. ezen a ponton egyértelmű, hogy a lokuszok távolsága és a rekombináció valószínűsége közötti arányosság elvész.
Az azonos kromoszómán társított lokusok „asszociációs csoportokat” alkotnak. A nagyon távoli lókuszok olyan valószínűséggel válhatnak szétválás útján, hogy egymástól függetlenül viselkednek, de mindegyik alacsonyabb rekombinációs sebességgel társul a köztes lókuszokhoz.
Amikor a társulási csoporton belüli sok génpár közötti rekombináció mértéke ismert, elkezdődhet a „genetikai térképek” felépítése. Figyelembe véve, hogy a két gén (a és b) közötti távolságot a rekombinációs arány fejezi ki, és hogy a távolság egy harmadik géntől c lehet akár az összeg, akár a b -től való távolság különbsége, lehetséges rekonstruálni a kölcsönös távolságok térképét, amely az adott asszociációs csoporton belül, vagyis annak a kromoszómának a genetikai térképe lesz.
Most általánosságban meg kell fontolnunk néhány fogalmat, amelyek korlátozzák a genotípusos karakterek fenotípusos megnyilvánulását.
Először is a penetrancia és az expresszivitás fogalmáról fogunk beszélni, majd különös figyelmet fordítunk a génhatás szabályozásának jelenségeire.
PENETRANCE
A gének penetrációja azt a képességet jelenti, hogy képes megnyilvánulni a fenotípusban. A behatolást statisztikailag mérik, számolva a fenotípusok gyakoriságát, amelyek azt mutatják, hogy a karakter az azt tartalmazó 100 genotípusból. A 0,7 penetranciájú tulajdonság olyan tulajdonság, amely genotípusos gyakoriságának 70% -ában fenotípusosan fordul elő.
Kifejezhetőség
Az expresszivitás a fenotípusos megnyilvánulás mértékének kvantitatív értékelése.
A GÉNEK AKCIÓJÁNAK SZABÁLYOZÁSA
A sejtek minden enzimet és fehérjét azonos sebességgel és időben állítanak elő. Az Escherichia coli sejteket például energia- és szénatomokkal lehet ellátni a laktóz-diszacharidból, mivel a béta-galaktozidáz enzimnek köszönhetően képesek glükózra és galaktózra bontani őket. körülbelül 3000 béta-galaktozidáz molekula van, ami az adott sejt fehérjék 3% -ának felel meg; laktóz hiányában csak egy molekula béta-galaktozidáz lesz baktériumsejtenként. A galaktozidáz új mRNS -molekulákból fog szintetizálódni, ha használható. Az enzimben gazdag E. coli mutáns törzsek akkor is ismertek, ha nincs laktóz: ezek a mutánsok hátrányos helyzetben vannak a normál sejtekkel szemben, mivel szükségtelen energia- és anyagfelhasználásra kényszerítik az enzim előállítását, amely szubsztrát nélkül marad. Azokat az anyagokat, amelyek az enzim mennyiségének növekedését okozzák, mint a laktóz esetében, induktoroknak, míg az enzimeket indukálhatónak nevezzük. Más anyagok is - bizonyos módon - bizonyos enzimek termelését idézik elő. Például az E. coliban, amely képes felépíteni minden aminosavát, szénnel és ammóniummal (NH3), egy adott aminosav (például hisztidin) jelenléte a tenyésztőközegben blokkolja a maga az aminosav bioszintézise: ezekről az enzimekről azt mondják, hogy represszívak. A baktériumsejtekben az mRNS -molekulákat nem sokkal a keletkezésük után lebontják, ezért az mRNS -termelés szabályozása az enzimatikus szintézis egyidejű szabályozását jelenti. idő.
AZ OPERON
Annak megmagyarázására, hogy a baktériumsejt hogyan tudja szabályozni saját enzimtermelését, Jacob és Monod megfogalmazták az operon hipotézisét; az operont számos gén alkotja, amelyek funkcionálisan rokonok és megszakítás nélkül illeszkednek a DNS -szakasz mentén Az operon három különböző típusú génből áll: a promoter, ahol az mRNS kialakulása kezdődik; az operátor, ahol az ellenőrzést gyakorolják; egy vagy több szerkezeti gén, amelyek enzimeket vagy más fehérjéket kódolnak. A béta-galaktozidáz rendszerben az operon a béta-galaktozidázon kívül két másik gént is tartalmaz, amelyek strukturálisan kódolják más enzimek, amelyek részt vesznek a laktóz metabolizmusában. Ezek a gének szomszédosak egymással, és ugyanazt a DNS -hélixet egymás után írják át egyetlen mRNS -molekulává.Az így előállított mRNS -molekulák nagyon rövid ideig aktívak, ezt követően specifikus enzimek elpusztítják őket.
Az operon aktivitását viszont egy másik gén, a szabályozó szabályozza, amely szintén távol lehet az operontól: ez a szabályozó olyan fehérjét kódol, amelyet elnyomónak neveznek, és amely úgy tűnik, hogy az operátor génhez kötődik a DNS -hez. a promoter és a szerkezeti gének között valójában gátolja az mRNS termelését.
Az elnyomót viszont irányítják, és a vezérlést "jelző" anyag segítségével hajtják végre. Az indukálható enzimek esetében ez az anyag az "induktor". Az indukálószer kötődik a represszormolekulához, megváltoztatva annak alakját, így már nem tud alkalmazkodni a DNS -hez: ebben az esetben, mivel nincs promotor és a szerkezeti gének között , a represszor képezheti az mRNS -molekulákat és ezekből a fehérjemolekulákat. Az induktor készletének kimerülése után a szabályozó visszanyeri az irányítást, ami leállítja az új mRNS, tehát az új fehérjék termelését. A béta-galaktozidáz rendszerben az induktor laktóz vagy ehhez nagyon hasonló anyag. Származéka: csatlakozik a represszorhoz, és inaktiválja azt, hogy lehetővé tegye az enzimek bioszintézisét. Represszálható enzimek esetében a "jelként" hatóanyag corepressorként működik: az represszor csak akkor aktív, ha a corepressorral kombinálják. A tucatnyi különböző enzimet tartalmazó hisztidin rendszerben ez az aminosav, a tRNS -jével, a corepressorral és a hisztidinnel kombinálva.
ALLOSZTERIKUS Kölcsönhatások
Az alloszterikus kölcsönhatások, amelyek magukban foglalják az enzim alakjának megváltoztatásával történő inaktiválását, más módon biztosítják a sejt metabolikus aktivitásának szabályozását. Az alloszterikus kölcsönhatások pontosabb szabályozást tesznek lehetővé, mint az operon induktor -represszor rendszere, de nem érik el azt a hasznos eredményt, hogy kizárják az adott anyag bioszintézisét az első szakaszból - az mRNS előállításából.
ELLENŐRZŐ RENDSZEREK AZ EUCARIUSBAN
Vannak olyan tények, amelyek arra engednek következtetni, hogy az operonhoz hasonló szabályozási rendszer működik és kiemelkedő a növények és állatok körében. Ezen organizmusok kromoszómái mélyen különböznek az E. coli és más prokariótákétól. A mitózis mechanizmusa olyan, hogy egy adott növény vagy állat minden sejtje rendelkezik minden információval
a megtermékenyített petesejtben jelen lévő genetika. Ezért bármely speciális sejtben a gének többsége a sejt élettartama alatt hatástalan marad. Ezekben a sejtekben a DNS mindig fehérjékhez kapcsolódik. Lehetséges tehát, hogy az eukariótákban a gének elnyomása pontosan ezt az összefüggést igényli a DNS és a fehérjék között.